
See
also: fungi, algae, cyanobacteria, epiphytes.
Lichens
are dual-organisms, consisting of an alga and a fungus (almost
always an ascomycete) living
together in symbiosis. Since fungi belong to
the fungal Kingdom, and algae to Protoctista, in the five
kingdom classification scheme, we have two organisms from
different kingdoms living in unity. In some
lichens the 'algal' partner, or photosymbiont (photobiont),
is a cyanobacterium, a prokaryote.
Cyanobacteria though once grouped together with algae are now
classified as prokaryotes, along with other bacteria; we can
think of them as prokaryotic algae. The algae lives within the
fungus body in symbiosis. Although often classified separately,
lichens are increasingly seen as fungi hosting algal symbionts.
The fungus can not live and grow without its algal partner and
is never found in nature without it. In contrast, the algal or
cyanobacterial partner is also found living free in nature.
The situation of the lichens is rather similar to that in
zoology, in many cnidarians (corals, hydra,
jellyfish, etc.) which have algae living within their bodies,
the cells of the algal partner being called
zoochlorellae in this case. In both cases the host benefits as
the algae can make food from
photosynthesis. Normally the algae would use all this food in
growth, but in these cases much of the
food produced is transferred to the host. In the jellyfish Cassiopea, the jellyfish swims
upside-down,
positioning itself in the light or resting on the bottom in
shallow water, exposing the algae, which are
situated in tissues on its underside, to optimal sunlight for
photosynthesis. The algae thus gets a
protective body (or 'house') and optimum sunlight and the
jellyfish gets food from the algae.
Similarly in lichens, the algae live in a protective fungal
body, which lifts them above the surface, and
thus closer to the light source than might otherwise be
possible, and the fungus derives food from the
algae. The benefit is more obvious for the fungus, though if one
considers the typical habitats of lichens
- tree trunks and branches, rocks (e.g. in the splash zone by
the sea, or in mountaineous terrain) then it
is apparent that the fungus elevates the algae above their
free-living competitors which remain on the
surface. This is good for obtaining light and carbon dioxide for
photosynthesis and also good for
dispersal. Lichens are also one of the few organisms that can
exist in the harsh dry stony Antarctic
deserts, in which case the fungal body undoubtedly provides
protection for the algal host. Many lichens
are also pigmented, and one function of these pigments is to
screen the algae from bright ultraviolet
light which may damage their photosynthetic apparatus.

Above:
a lichen seen in section. Many, but by no means all, lichens
have a layered or stratified thallus,
like that shown above. The body or thallus of the lichen is
composed of elongated multinucleate
threads (rather than 'conventional' cells) called hyphae, typical of fungi.
(Though the hyphae may
have cross-walls to divide them into cellular units, as in
lichens, so we can speak about 'fungal cells').
The algae tend to be concentrated in a layer in the upper
medulla (and sometimes also the inner
layers of the cortex) of stratified lichens. The medullary
hyphae are intimately associated with their
algal partner in various ways. The hyphae of the cortical
regions are more-or-less aligned, and so
appear cell-like as pseudoparenchyma in section. The hyphae
of the cortical zones are typically
thick-walled and glued together by materials secreted by the
hyphae (not shown in our model), filling
the spaces between the hyphae and forming the so-called conglutinate zone. These gluing together
of the hyphae not only gives the fungal body strength, but helps
protect it from adverse conditions.
The hyphae of the medulla are more loosely packed and not
cemented together, instead air-spaces
occur between them, allowing the easy delivery of carbon-dioxide
to the algal symbionts (this is similar
to the arrangement of plane cells in the spongy mesophyll of a leaf).
The conglutinate layer makes it hard for enough oxygen for
photosynthesis to diffuse into the medulla,
to counter this lichens often have pores in the undersurface of
the thallus - orderly breaks in the
cortex, allowing gases to diffuse in and out of the medulla
directly. These pores have a different
structure to the stomata of plants and are called cyphellae (singular cyphella),
which have a cellular
structure, or pseudocyphellae if they are simple
pores or breaks in the cortex, each of which may be
borne on a small wart-like structure. These pores allow gas
exchange, taking in oxygen for the fungus
and carbon dioxide for the photosymbiont. Respiration of the
fungal host also provides carbon dioxide
for photosynthesis. The algal partner constitutes up to about
20% of the mass of a typical stratified
lichen thallus. About 30-50% of the volume is taken up by the
medullary air-spaces.
Some lichens have a covering of dead fungal cells/hyphae on the
thallus surface, called the pruina.
Some have a covering of salt crystals or hairs. A felty covering
of hyphae, on the lower or upper
surface of a thallus, is called a tomentum. In many lichens,
growth of the fungal hyphae at the margin
of the thallus, forms a black border, called the prothallus.

Cyanobacterial
partners
The
lichen Peltigera (lichens are named
after the fungal partner) associates with the cyanobacterium
Nostoc. Microcolonies or
groups of cells of Nostoc, encased in a common
gelatinous envelope of their
own secretion, occur within the fungus body. Side-branches from
the medullary hyphae penetrate the
gelatinous capsules of the Nostoc microcolonies, but do
not contact or penetrate the Nostoc cells
themselves. These hyphal branches or projections have simplified
cell-walls, and are thus not
themselves hyphae. Presumable the lack of certain fungal
hyphal-wall layers in these projections
enables them to more rapidly exchange materials between the
fungal and algal partners. The Nostoc
cells are also seen to bud off extracellular outer
membrane vesicles
(OMVs - a characteristic of
certain Gram-negative bacteria like Nostoc and Bacilus
fragilis,
which form by budding from the outer
membrane, an outer layer of the
cell envelope). These may also be involved in transport or
communication between the cyanobacterial and fungal partners.
Lichens are able to tolerate periodic
dehydration and Peltigera requires contact with
liquid water to rehydrate it sufficiently for Nostoc to
begin photosynthesising. Cyanobacterial symbionts have a further
advantage - they may fix
nitrogen
for the lichen.
Green
eukaryotic algal partners
The
single-celled alga Coccomyxa forms lichen
associations with ascomycete and basidiomycete
lichens. These lichens can rehydrate sufficiently in moist air
for phorosynthesis to begin. Fungal
hyphae are in intimate contact with the algal cells, the two
being fused together along part of their
length, with the hypha penetrating the outermost layer of the Coccomyxa cell wall/capsule.
Certain 'leafy' (foliose) and 'shrub-like' (fruticose) lichens form
symbioses with the green
single-celled alga Trebouxia (in total some 40% of
lichens associate with this alga). The Parmeliaceae
comprise an ascomycete family of such lichens. In this family
sexual reproduction of the fungus host
tends to be reduced, reproduction predominantly occurring
asexually by releasing propagules which
contain both fungal hyphae and algal cells, which reduces the
problem of a young fungus acquiring its
algal host before growth can resume. The hyphae give off
projections, called haustoria, that form tight
contacts with the algal cells, allowing the direct exchange of
materials between the fungus and algal
partners. In these types, at least, fixed-carbon containing
materials (such as polyols or sugar-alcohols
like sorbitol (hexan-1,2,3,4,5,6-hexol)) produced by the algae
in photosynthesis are secreted and are
likely transported along the cell wall of the haustoria and
hypha, which are in intimate contact with the
algal cell wall - there is apoplastic
continuity
between the alga and the fungus (see transport in
plants).

Above:
a fruticose lichen (centre) and encrusting foliose lichens
(right) growing on a branch in a
mixed oak woodland.
Some
lichens can partner with either a cyanobacterial partner or a
eukaryotic alga. The morphology of the lichen may be very
different depending which alga is present, resulting in
different species names being assigned. Sometimes a chimaeric
thallus may result, with both morphologies present in different
regions of the same thallus, depending on the alga present.
Some lichens make use of the nitrogen-fixing properties of
cyanobacteria by forming specialised
structures containing them, called cephalodia (singular cephalodium)
which may be spherical blebs of tissue borne on the thallus
surface, or may be hidden within the thallus, the rest of the
thallus containing a eukaryotic algal symbiont. Others have a
layer of cyanobacteria and a layer of eukaryotic algae within
the same tissue.
Various root-like structures, called rhizinae, may extend from the
bottom of the thallus, which serve primarily for anchorage
rather than the absorption of water. These may be simple hairs,
bulbous structures, bearing whorls of side-branches, branch by
forking, or highly-branched fibrous structures. Some lichens
have very tough thalli and are very well anchored to the
substrate on which they grow. An example are the encrusting
lichens of the splash zone, growing on rocks above the high tide
line. Exposed to the elements these lichens are extremely tough
and very hard to remove and may be mistaken for tar.
Reproduction
of Lichens
Many
lichens reproduce sexually by the normal means of the fungus
host. This sexual reproduction involves only the fungal partner.
Ascomycete lichens, for example, form characteristic ascomycete
sporing structures (sporocarps): cup-like structures (apothecia) or pear-shaped flasks
(called perithecia) open at one end. The
spores germinate, but must contact the correct algal partner
(which is free-living) quickly or else they die. Rarely algae
may be ejected in packages with the sexual spores. The inner
lining of these sporocarps is called the hymenium, and consists
of sexual spore-containing fleshy hair-like structures called asci, and sterile hairs
called paraphyses. The sexual spores, or
ascospores, are fired from the
asci as they rupture when ripe.
To counter these problems, many lichens reproduce primarily by
asexual means. Asexual spores,
called soredia (singular soredium),
consisting of a few algal cells wrapped in fungal hyphae, are
released from the medulla through cracks in the cortex. Regions
of the lichen thallus, called soralia
(singular
soralium) may be present - these are regions specialised for
soredia production. The soralia may be diffuse regions with no
definite structure, or they may be delimted or marked off as
distinct regions or structures at the tips of the thallus or on
its surface; these may be of various shapes and are often
distinct protuberances or patches.
Sometimes protuberances, which maybe branched, called isidia (singular isidium),
develop, which have lines of breakage built in at the base (the
cortex is absent in these breakage points, so that only the weak
medulla holds the protuberance to the thallus). These
protuberances differ from soredia in that they are surrounded by
cortex. The soredia may form a grey powder that covers the
surface of the lichen and easily rubs off to the touch.
Sometimes the soralia/soredia form by the breaking off of
isidial tips, in which case they are called isidial
soralia.
Some lichens produce minute pear-shaped structures inside the
thallus, which produce spores called conidia, which are presumed to
be asexual spores.
Some lichens occur in species
pairs.
These are 'species' with similar chemistry and form, but in
which one is always fertile and does not reproduce asexually
whilst the other rarely reproduces sexually but produces asexual
soredia and/or isidia.
Many lichens can resist periodic dehydration, becoming dormant,
dry and brittle when dehydrated, and rapidly re-hydrating and
resuming metabolism and growth when moist. Dry lichens may
easily fragment, each fragment regenerating into a new lichen
when moisture returns.
Most lichens are slow growing and reach only a few centimetres
in width, though some tropical fruticose forms hanging from
trees, reach several metres in length. Many grow only 1
millimetre a year. Fruticose lichens may live ten years or more,
but encrusting types may live for centuries. By measuring their
diameter on dated tombstones in any locale, it is possible to
estimate their rate of growth. Foliose and fruticose forms grow
faster, e.g. Peltigera can grow 2-3 cm a
year.
Habitats
Lichens
often fluorish in extreme environments, from mountains, to under
the water along the seacoast, to tropical and Antarctic deserts.
In particular they can rapidly dehydrate, without suffering
damage, entering a dormant state in which the cellular machinery
is protected from adverse conditions; and then they can rapidly
rehydrate and resume their function when moisture returns.
However, they can not grow when dry and their inability to
conserve water means that most species favour humid conditions.
Lichens are often pioneers. This means that they
are among the first to colonise certain newly formed habitats.
In particular, freshly-exposed rocks are a good surface for
colonisation by lichens (and bacteria and cyanobacteria). This
may result from a land slide, other forms of erosion, or from
volcanic, alluvial or glacial deposition, for example weatehring
limestone and granite formations and eroding coastlines. Such a
surface favours the encrusting or crustose lichens - lichens
whose thallus is tightly pressed to the stony surface and very
hard to remove. This is beneficial as these habitats are often
exposed mountaineous or coastal habitats. It is remarkable how
tightly some of these lichens anchor to the rock surface. Along
with bacteria who oxidise rocks and the natural effects of
weathering, such as freeze-thaw cycles that tend to shatter
rocks, lichens contribute to the slow process of soil formation.
In time, as soil forms, plants may colonise the area. In this
way the dominant vegetation may change over time in a series or
succession. In such cases lichens
may remain, as in woodlands, though not as the dominant flora.
However, in the harshest of habitats, such as at very high
altitudes, along exposed rocky coastlines and in the Antarctic
deserts, the lichens may remain the dominant flora and form the
climax vegetation.
Lichens also act as important reservoirs of water. They rapidly
hydrate when wet, absorbing water that drips down tree trunks or
strikes the soil surface, acting like sponges. This helps reduce
flooding by reducing the run-off of rainwater across the soil
surface, and instead delays the arrival of that water into
streams and rivers, allowing time for excess water to drain.
Lichens
as pollution indicators
Many
lichen species are very susceptible to industrial air pollution,
whilst others are more resistant. The mean level of sulphur
dioxide gas, produced largely anthropegenically by burning
fossil fuels, can be estimated by analysing the growth of
lichens on trees. If a tree only has green algae growing on its
base and no lichens, then levels are very high (over 170
micrograms per cubic metre of air). In slightly lower
concentrations of sulphur dioxide, green algae may grow along
the whole height of the trunk whilst the grey-green crustaceous
(encrusting or crustose) lichen Lecanora may be present, either
at the base only, or along the height of the trunk, depending on
pollution levels. Lecanora prefers high sulphur
dioxide levels and is absent in cleaner air (at least in Europe)
and was unknown in Europe before 1860. In Europe it is an urban
lichen. Fruticose and foliaceous species, such as Parmelia and Lobaria occur where pollution
levels are very low or natural. Humidity is also a factor in
determining the extent of coverage of tree bark by lichens.
It is remarkable how often in evolution the greatest advances
occur through cooperation, even
between different species, rather than by blatant and direct
competition. Concepts such as these have modified and extended
the Darwinian notion of 'survival of the fittest' and form part
of the modern neo-Darwinian theory of evolution. It is also
remarkable that lichens have evolved into leaf-like structures
with similar shapes and internal morphology, though this is
perhaps not surprising since both are subject to the same
physical constraints. This is an example of convergent evolution
- similar structures evolving independently and in quite
different ways.
Further
Reading
The
following is a good paper on the algal-fungal symbiosis,
focusing on the structure of the contacts between them and the
mechanism of material transfer between the two symbionts:
R. Honegger, 1991. Functional aspects of the lichen symbiosis. Annu. Rev. Plant
Physiol. Plant Mol. Biol.
42: 553-578.

Above:
Cup-like apothecia of a lichen growing on a stone post in a
wood. This is probably
Xanthoria (probably Xanthoria parietina) although Caloplaca and Fulgensia are similar
and best distinguished by microscopical examination. Below: Xanthoria parietina growing
on an elder (Sambucus
nigra)
twig.

Was
this organic growth an inspiration for the Zygons?
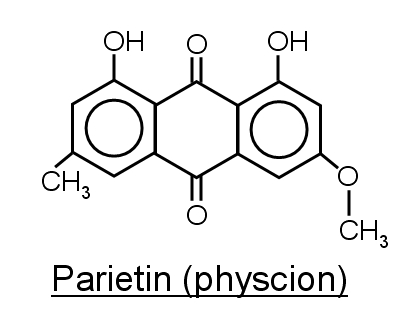
The
orange pigment, parietin (also called physcion) is produced
by some lichens like Xanthoria
parietina.
This pigment protects
the fungus from damaging UV radiation and also has antifungal
properties. It is also a potential anti-cancer agent.
| Lichens |
| |
|
|
|
|
|