| Insect Nutrition |




| | | | | | | | | |
| | | | | | | | |
Insect nutrition is extremely diverse, and the mouthparts, gut structure and mode of nutrition vary enormously,
however the gut can be divided into the following regions: foregut, midgut and hindgut. The lining of the of the
foregut and midgut is continuous with the epidermis (skin) of the insect and are lined by cuticle (which must be
shed during molting). We shall look at each structure in turn, beginning with the mouthparts.
however the gut can be divided into the following regions: foregut, midgut and hindgut. The lining of the of the
foregut and midgut is continuous with the epidermis (skin) of the insect and are lined by cuticle (which must be
shed during molting). We shall look at each structure in turn, beginning with the mouthparts.
Insect mouthparts are extremely diverse, arguably coming in more different forms and with more different modes of
operation than any other organ-system in the animal kingdom. The most unspecialised form (from which the others
are thought to be derived) are crushing and slicing jaws or biting mouthparts. A good example is the
grasshopper and locust. Grasshoppers use their jaws to slice and crush plant leaves. The grasshopper has
hypognathus mouthparts, meaning they point downwards, as it stands over its food when eating. (Rove beetles,
such as Aleochara, also have biting mouthparts, but these are prognathus, pointing forwards as this beetle uses
its jaws to tackle prey items, insect larvae in the case of Aleochara). A third form are opsithognathus mouthparts
in which the head is tucked underneath the body with the mouthparts projected backwards.
The mouthparts consist of three pairs of appendages (that are developmentally and evolutionary legs that have
become modified, there being one pair of limbs per body segment, with several segments compacted together to
form the head, see animal bodies). The furthest forward pair are the mandibles, unsegmented pincer-like jaws
that operate from side-to-side. They are covered in thick hard cuticle (they are highly sclerotised) and are hinged
to the head at two points. The cuticle of the head has a thickened ridge to strengthen it where the mandibles
attach (this ridge corresponds to the furrow, visible externally, between the frons and clypeus, major chitinous
plates that form the exoskeleton of the head). Large fan-shaped muscles operate the madibles, inserting at the
hinges and fanning out to insert on the cuticle at the top of the head. Smaller muscles give the mandibles fine
movements.
operation than any other organ-system in the animal kingdom. The most unspecialised form (from which the others
are thought to be derived) are crushing and slicing jaws or biting mouthparts. A good example is the
grasshopper and locust. Grasshoppers use their jaws to slice and crush plant leaves. The grasshopper has
hypognathus mouthparts, meaning they point downwards, as it stands over its food when eating. (Rove beetles,
such as Aleochara, also have biting mouthparts, but these are prognathus, pointing forwards as this beetle uses
its jaws to tackle prey items, insect larvae in the case of Aleochara). A third form are opsithognathus mouthparts
in which the head is tucked underneath the body with the mouthparts projected backwards.
The mouthparts consist of three pairs of appendages (that are developmentally and evolutionary legs that have
become modified, there being one pair of limbs per body segment, with several segments compacted together to
form the head, see animal bodies). The furthest forward pair are the mandibles, unsegmented pincer-like jaws
that operate from side-to-side. They are covered in thick hard cuticle (they are highly sclerotised) and are hinged
to the head at two points. The cuticle of the head has a thickened ridge to strengthen it where the mandibles
attach (this ridge corresponds to the furrow, visible externally, between the frons and clypeus, major chitinous
plates that form the exoskeleton of the head). Large fan-shaped muscles operate the madibles, inserting at the
hinges and fanning out to insert on the cuticle at the top of the head. Smaller muscles give the mandibles fine
movements.
The jointed maxillae are a pair of secondary jaws, behind the mandibles, and which also operate from
side-to-side. These manipulate food and guide it towards the mouth where it is crushed by the
mandibles.
The labium, or lower lip, is actually a fused pair of secondary maxillae. Both the maxillae and the labium
bear a pair of segmented appendages, the palps (one pair of maxillary palps are attached to the
maxillae, and one pair to the labium, resulting in four palps in total). The palps are sensory, bearing
taste sensors (gustatory sensilla). The flaplike labrum is the upper lip. The mouth is situated between
the labrum and labium at the base of the mandibles and opens into the buccal cavity, the first
compartment of the foregut. The anterior surface of the labium bears the 'tongue' or hypopharynx
(external to the mouth in insects) which helps push food into the mouth.
These basic mouthparts are greatly modified in many insects. Many flies have sucking mouthparts,
which draw up liquids. These insects have a beak or proboscis for sucking up liquids. Many flies will
'spit' digestive enzymes onto their food and then suck up the liquefied partially-digested food. In bees,
the maxillae and labium are elongated into a tube: fused at their bases with the ends fitting together into
a tube for sucking up nectar. A joint between the fused base and the rest of the proboscis allows the
bee to fold away its proboscis under its head when not in use. In bees, wasps and ants, the mandibles
remain for digging and nest construction, although feeding is by sucking mouthparts.
In the fly proboscis (see figure below), which consists of elongated labrim, labium and hypopharynx
fitted together, saliva containing digestive enzymes is conveyed by a salivary channel in the
elongated hypopharynx and 'spat' onto the food. The partially digested liquid food is then sucked up the
food channel formed between the labium and labrum. The mandibles are absent and the maxillae
present as maxillary palps only. The main body of the proboscis is formed by the labium and the two
lobes at the end of the proboscis are modified labial palps, with ridged furrows underneath which
channels the liquid food into the food channel of the proboscis. The proboscis is jointed and can be
neatly folded away when not in use.
side-to-side. These manipulate food and guide it towards the mouth where it is crushed by the
mandibles.
The labium, or lower lip, is actually a fused pair of secondary maxillae. Both the maxillae and the labium
bear a pair of segmented appendages, the palps (one pair of maxillary palps are attached to the
maxillae, and one pair to the labium, resulting in four palps in total). The palps are sensory, bearing
taste sensors (gustatory sensilla). The flaplike labrum is the upper lip. The mouth is situated between
the labrum and labium at the base of the mandibles and opens into the buccal cavity, the first
compartment of the foregut. The anterior surface of the labium bears the 'tongue' or hypopharynx
(external to the mouth in insects) which helps push food into the mouth.
These basic mouthparts are greatly modified in many insects. Many flies have sucking mouthparts,
which draw up liquids. These insects have a beak or proboscis for sucking up liquids. Many flies will
'spit' digestive enzymes onto their food and then suck up the liquefied partially-digested food. In bees,
the maxillae and labium are elongated into a tube: fused at their bases with the ends fitting together into
a tube for sucking up nectar. A joint between the fused base and the rest of the proboscis allows the
bee to fold away its proboscis under its head when not in use. In bees, wasps and ants, the mandibles
remain for digging and nest construction, although feeding is by sucking mouthparts.
In the fly proboscis (see figure below), which consists of elongated labrim, labium and hypopharynx
fitted together, saliva containing digestive enzymes is conveyed by a salivary channel in the
elongated hypopharynx and 'spat' onto the food. The partially digested liquid food is then sucked up the
food channel formed between the labium and labrum. The mandibles are absent and the maxillae
present as maxillary palps only. The main body of the proboscis is formed by the labium and the two
lobes at the end of the proboscis are modified labial palps, with ridged furrows underneath which
channels the liquid food into the food channel of the proboscis. The proboscis is jointed and can be
neatly folded away when not in use.
Above: the head and sucking mouthparts of the fruitfly Drosophila. A: arista (part of the
antennae); F, funiculus (part of the antenna); L, labium ending in one pair of lobe-like labial
palps bearing furrows; P, proboscis, consisting mainly of the labium. Bottom right: transverse
section through the proboscis of the housefly, Musca.
antennae); F, funiculus (part of the antenna); L, labium ending in one pair of lobe-like labial
palps bearing furrows; P, proboscis, consisting mainly of the labium. Bottom right: transverse
section through the proboscis of the housefly, Musca.

mouthparts. The proboscis is modified to form a group of hard sclerotised lances, called stylets, for
puncturing the animal or plant. In the mosquito, a biting fly, for example, the labrum, maxillae and mandibles
are greatly elongated and lance-like and fit together into a single tubular set of stylets enclosed by the
labium. When it lands on a host, the mosquito probes the host's skin with the tip of its labium, which bears
taste sensors and other sensors. The mandibles and maxillae then stab alternately, piercing the skin. If the
stylet reaches its target, a small blood capillary, then being flexible it may extend part-way along the inside
of the capillary. The slivary channel of the hypopharynx conveys saliva into the wound, preventing blood
coagulation and beginning digestion with the enzymes it contains. (A host reaction to the salivary fluids
produces the itchy red lump of the mosquito bite). When the gut is full of blood and distended then it
signals the mosquito's brain to end feeding.
puncturing the animal or plant. In the mosquito, a biting fly, for example, the labrum, maxillae and mandibles
are greatly elongated and lance-like and fit together into a single tubular set of stylets enclosed by the
labium. When it lands on a host, the mosquito probes the host's skin with the tip of its labium, which bears
taste sensors and other sensors. The mandibles and maxillae then stab alternately, piercing the skin. If the
stylet reaches its target, a small blood capillary, then being flexible it may extend part-way along the inside
of the capillary. The slivary channel of the hypopharynx conveys saliva into the wound, preventing blood
coagulation and beginning digestion with the enzymes it contains. (A host reaction to the salivary fluids
produces the itchy red lump of the mosquito bite). When the gut is full of blood and distended then it
signals the mosquito's brain to end feeding.
True bugs (order Hemiptera, the hemipterans) and their relatives include many parasites of plants, such as
aphids, leafhoppers, cicadas and scale insects. The labium forms a beak-like sheath for the stylets. There
are four stylets, one pair of mandibles and one pair of maxillae, that fit together so as to form two internal
channels - a food channel and a salivary channel. Again, when feeding the labium folds back to expose the
stylets and alternate movements of the stylets puncture the host. Many bugs also feed upon animals,
including in some species other arthropods (e.g. assassin bugs) upon which they prey, sucking out the body
fluids.
aphids, leafhoppers, cicadas and scale insects. The labium forms a beak-like sheath for the stylets. There
are four stylets, one pair of mandibles and one pair of maxillae, that fit together so as to form two internal
channels - a food channel and a salivary channel. Again, when feeding the labium folds back to expose the
stylets and alternate movements of the stylets puncture the host. Many bugs also feed upon animals,
including in some species other arthropods (e.g. assassin bugs) upon which they prey, sucking out the body
fluids.
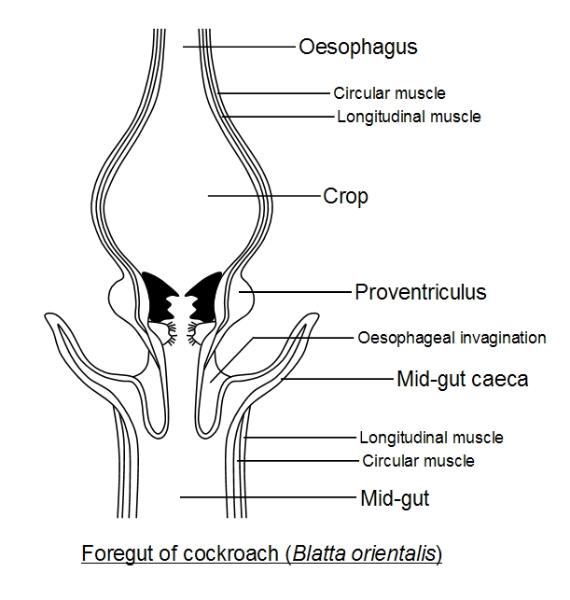
The insect Gut - Foregut
The mouth opens into the first chamber of the foregut, the buccal cavity, which leads into the tubular
storage space for food awaiting processing. Part of the foregut may also be modified to form a proventriculus,
where the cuticle lining the foregut may be modified into crushing and grinding teeth. It is here that chewing
The mouth opens into the first chamber of the foregut, the may take place. A circular muscular valves, the
cardiac sphincter, which leads into the tubular, separates the foregut from the midgut. buccal The
epithelium lining the forgut may continue past the sphincter, projecting partway into the midgut as the The
epithelium lining the forgut may continue past the sphincter, projecting partway into the midgut as the
oesophageal fold.
The mouth opens into the first chamber of the foregut, the buccal cavity, which leads into the tubular
storage space for food awaiting processing. Part of the foregut may also be modified to form a proventriculus,
where the cuticle lining the foregut may be modified into crushing and grinding teeth. It is here that chewing
The mouth opens into the first chamber of the foregut, the may take place. A circular muscular valves, the
cardiac sphincter, which leads into the tubular, separates the foregut from the midgut. buccal The
epithelium lining the forgut may continue past the sphincter, projecting partway into the midgut as the The
epithelium lining the forgut may continue past the sphincter, projecting partway into the midgut as the
oesophageal fold.
Function of the Crop
This acts as a temporary food store when the midgut is filled to capacity. It stores mostly liquid food, whether
blood sucked up by a tsetse-fly (Glossina) or partially-digested food sucked up by a housefly (Musca). In the
mosquito, blood goes straight to the midgut, whilst fruit juices are stored in the crop (the labium has sweet
taste-sensors, the labrum blood taste-sensors). In the cockroach, Periplaneta, some digestion occurs in the crop
and here fatty acids (from the digestion of fats) are absorbed. The crop may contain still active salivary enzymes
and also enzymes refluxed forwards from the midgut. In bees, nectar is converted into honey, whilst in the crop,
by salivary enzymes.
The crop also has a function during molting in some insects, taking up air to expand the insect during shedding of
the cuticle and during enlargement of the insect prior to the new cuticle hardening.
Oesophageal Diverticula
Blind ending tubes or sacs may extend from the oesophagus; these are called diverticula. In saw-fly larvae
(Lophyrus) a pair of such pouches receives useless resin from the surface of pine needles on which it feeds. In
mosquitoes the diverticula take up any air sucked up with the food.
Function of the Proventriculus
This may simply be a sphincter or a muscular organ armed with spines or teeth, also ending in a sphincter. It
controls the passage of food into the midgut, acting like a sieve. In the cockroach, Periplaneta, a ring of six
powerful teeth break-up and compact the food. Behind these are two rows of hairy cushions. In the bee, the crop
filters out pollen grains that are passed onto the midgut, whilst retaining fluids such as nectar.
The Midgut
The mid-gut has muscular walls and is lined on the inside by cubical or columnar epithelium bearing microvilli or
some similar structure to increase the surface area for the exchange of materials with the gut contents. The
midgut lumen is not lined by cuticle, but is covered by the peritrophic membrane which protects the epithelium
and is secreted by the epithelium. This membrane may contain chitin, proteins and glycogen and is moulded by
elaborate mechanisms under the control of the gut wall musculature. It is periodically shed (and regenerated) and
encloses undigested food as it passes into the hindgut, forming a sheath for the faecal pellets.
The midgut secretes digestive enzymes and also absorbs many digested nutrients, such as glucose (the product
of carbohydrate digestion) and amino acids (the product of protein and polypeptide digestion). The fat body of
the insect converts absorbed glucose into trehalose, the sugar found in insect 'blood' (haemolymph) and body
fluids. Transporting sugar around the body in the form of trehalose makes it much harder for bacteria to
metabolise and this helps limit infection. (In mammals, the nervous system requires glucose and carrying glucose
in the warm blood necessitates a very advanced immune system in mammals). The rehalose is converted into
glucose for use when taken up by the cells.
The midgut may have distinct subdivisions. For example, in the tsetse fly (Glossina) the first region absorbs water
from the blood meal; the second region releases enzymes (causing the blood to blacken as it is digested); and
the final region absorbs the products of digestion. In some Homoptera, which feed on plant juices, the gut folds
around so that the oesophagus opens both into the front of the midgut, as usual, and also into a special filter
chamber in the back of the midgut. This allows excess fluid to short-circuit the digestive action of the midgut to
be excreted (the Malpighian tubules cover the surface of the filter chamber). Pouches or midgut caecae may
extend from the midgut (as in Aleochara at the top of the page) increasing its surface area for release of enzymes
or for absorption.
The Hindgut
A circular muscular valve, the pyloric sphincter, separates the midgut from the hindgut. The hindgut is lined by
cuticle often bearing tiny backward-pointing spines to help draw the faecal pellet backwards. The faecal pellet is
encloded in the shed peritrophic membrane of the midgut. The rear of the hindgut may form a distinct rectum.
This may bear six longitudinal folds or rectal pads or glands that may allow the rectum to expand when needed
and probably absorb water (the rectal pads are richly supplied by oxygen via tracheae and so are metabolically
active and the cuticle covering them is often thin and even porous). The rectum thus appears to act to conserve
water by absorbing water from the faeces. When you think how little water there is in a small insect, it is
remarkable how they can survive for so long between meals without drying up! A starved fly is thought to typically
run out of fuel (glucose/trehalose) before it dies by dehydration.
The faecal matter becomes progessively drier as it passes along the hindgut and ammonia and bicarbonate may
be secreted into it as the hindgut appears to regulate the pH (acidity) of the insect body. In the cockroach food
takes 9-33 hours to pass through the gut, in the silkworm 2-3 hours. Faeces are expelled from the rectum through
the anus, typically as dry pellets, except in those insects that have excess fluid in their diet, such as plant
sap-sucking Hemiptera, flies (Diptera), nectar-feeding moths and butterflies (Lepidoptera) and Hymenoptera
(bees, ants and wasps). In aphids and coccids (scale insects) it contains excess carbohydrates from the sugary
phloem sap of the host plant and is expelled as droplets of 'honey-dew' (often coated in wax in aphids to prevent
the aphid colony becoming covered in sticky honey-dew!). The beetle Platydema spins a pupal cocoon during
metamorphosis of chitinous fragments glued together, derived from the fungus on which it feeds.
Dietary Requirements and Symbionts
Insects are truly remarkable in their abilities to utilise unlikely food sources. They are also remarkable for the
efficiency with each they feed and grow, often on a tiny amount of food. Insects with distinct larval stages also
have the advantage of utilising different food sources, with the larvae avoiding competition with the adults by
eating very different food substances, such as the hungry caterpillar who eats foliage whereas the adult moth or
butterfly may feed on nectar. In many cases the larvae are aquatic and so feed in a very different environment to
the terrestrial adults. Almost anything organic is a food source for at least one species of insect. Cockroaches
can live on paper and a little water, silverfish may be kept on nothing more than a small piece of potato. House
flies, Musca domestica, can be grown using meat for their larvae (on which they will lay their eggs) and milk
powder and perhaps a little cholesterol, and water for the adults. The rove beetle, Aleochara bilineata, has a
high-protein diet of fly maggots, but can be reared happily on cat biscuits!
A good example to consider are the wood-boring insects. An example is Lyctus, the powderpost beetle, whose
larvae bore into wood, leaving behind powdery frass (faeces) of undigested wood. Wood is typically about 50%
cellulose ( a carbohydrate), 40% lignin and 10% hemicelluloses (another carbohydrate) and small amounts of
starch and other carbohydrates. Lyctus feeds on the latter - the starch and other sugars, but cannot digest the
main cellulose and lignin bulk of the wood which passes through the gut to be defecated as the powder which
gives the insect its common name.
Termites, however, have flagellated pro digest particles of wood and break-down the cellulose. The insect can
then digest excess protozoa and so derive nourishment. In this way termites will extract much of the cellulose from
ingested wood, but none of the lignin. Mixotricha is one of these protozoal endosymbionts found in the gut of
the termite Macrotermes (endosymbiont - an organism that lives inside another organism) and is a very unusual
single-celled organism indeed! Mixotricha uses its four flagella to steer when swimming, but locomotive power is
achieved by hundreds of thousands of spirochaete bacteria that live attached to the outside of the protozoan cell,
functioning as cilia, and drive it along (by means of their endoflagella). Spherical bacteria living inside the
protozoan cell provide it with energy by respiration, functioning as mitochondria which it lacks. Crucially, for the
termite, Mixotricha has at least two bacterial endosymbionts that can produce cellulase, the enzyme which
degrades cellulose into smaller carbohydrates, allowing it to degrade the particles of wood that it ingests. Thus,
the termite is a complex example of different organisms living together as a single unit! No animal is known to
synthesise its own cellulase, but to rely on bacteria to synthesise it for them. Cellulase is a bacterial invention! No
insect appears able to digest lignin, and this is defecated. (Some bacteria and fungi do digest lignin, often at a
very slow rate - lignin really is tough stuff!).
Many insects, if not all, rely on bacteria and other microorganisms living within their guts to assist in food
digestion. Vertebrates have a similar dependency; in herbivorous mammals bacteria in the gut secrete cellulose
to help digest plant cell walls and these live within special fermentation chambers, such as the appendix and
large intestine or a special additional compartment of the stomach, as in ruminants. The guts of many insects also
have special fermentation chambers. These chambers may be extensions or caecae of the midgut, such as the
bacterial crypts of Aphanus, which feeds on plant juices; or there may be a special chamber in the hindgut, such
as the rectal pouch of the termite Eutermes, which contains flagellates, or the fermentation chamber of the
lamellicorn larva. In humans the bacteria inside in the gut lumen, as they do in many insects, but in many insects
the symbionts are often intracellular - living inside the cells, often special cells in the gut lining.
Humans acquire their bacteria from the environment and from their diet, but in many insects the microorganisms
are very specific and it may be crucial that the insect acquires its special bacterial symbionts. In these cases the
mother often passes them on to the offspring. In the bed bug Cimex, the symbionts enter nurse cells which
surround and nourish the developing egg in the ovary and pass from here into the egg cell itself. In a similar
manner rod-shaped bacteria living in the fat body of the termite Macrotermes are transmitted to the egg cell whilst
in the ovary. A similar invasion of the egg, but just before the egg shell is secreted, occurs in many insects, and
Lyctus (powderpost beetle) mentioned earlier. In some insects the bacteria infect the male reproductive system
and are then passed onto the male and enter the egg through the micropyle before it is laid (the micropyle is a
pore in the shell through which the sperm enters during fertilisation). There may be special glands attached to the
male reproductive system that store and release the microbes for this purpose. In some the female contaminates
the eggs when they are laid and the hatchlings eat all or part of the egg-shell, or probe its surface, to pick up
their microbial partners. The symbionts may be bacteria, protozoa or fungi.
Other uses of the mouthparts
Some insects have evolved diverse uses for their mouthparts. Ants are a good example and the behaviour
patterns of Formica fusca have been well studied by D. I. Wlllis (Behavioural patterns of the ant, Formica fusca,
1962. Animal Behaviour 10: 105-111). The ant Formic fusca feeds on liquid foods by lapping with its 'tongue': a
bulbous projection, properly called the hypopharynx, on the labium (lower lip). During this ingestion of fluids the
mandibles are closed and maxillary palps are held back out of the way. The antennae are held in a W shape with
their tips resting on the surface of the food along with the labial palps - both antennae and labial palps seem to
be involved in tasting the food.
These ants also use their mouthparts for cleaning. When cleaning the antennae and forelegs, the fore-part of the
body is lifted 30 degrees above the horizontal, so that the middle and hindlegs are left supporting the body. Left
and right appendages are cleaned alternately. For example, when cleaning the right antenna, the right foreleg is
wiped down the antenna which is drawn through the strigil (a comblike spur of more-or-less rigid hairs) of the
foreleg as the head is held inclined towards the side being cleaned. The antenna is wiped one to three times in
succession whilst simultaneously the left foreleg is drawn through the mouthparts on the left side, between the
maxilla and labium to be cleaned by the maxillary comb (and possibly the hypopharynx). Sometimes, as part of the
same sweeping movement, the right foreleg may begin by sweeping over the back of the head and eye to clean
these structures too. The mandibles may also be brushed downwards by the left foreleg before it is drawn
through the mouthparts. This pattern is reversed as the left antenna and right foreleg are cleaned and this
alternating pattern is repeated several times.
An alternative posture is adopted when cleaning the middle and hindlegs. This time the ant leans away from the
side being cleaned and raises the legs on the side being cleaned whilst its body is supported on the middle and
hindlegs of the opposite side and the abdomen (forming a stable tripod). When cleaning the left legs, for example,
the left hindleg is cleaned by the left midleg brushing down it (which maybe assisted by the forelegs). The left
midleg is then cleaned by both forelegs brushing down it in succession. The tarsi ('feet') are cleaned mainly. Thus
there is a sequence in which foreign particles are passed from the hindlegs to the middle legs to the forelegs and
then to the mouthparts. Forelegs may also be brushed down each other as one leg is pulled through the strigil of
the other. Hindlegs may also brush the side of the abdomen before being cleaned.
Abdominal cleaning utilises a third posture. The body is raised and held clear of the ground by all six legs as the
head bends underneath the thorax whilst the abdomen is bent forwards towards the head. The mouthparts then
lick the end of the abdomen. The ants may also lick one-another, coating each other with saliva and cleaning
parts that the ant cannot reach by itself. Mouthparts, the abdomen and limb joints are especially focused on. The
ant being cleaned remains passive as the cleaning ant plays its antennae over the surface of the recipient.
Wallis speculated that the oily saliva of the ant may contain anti-microbial properties, as well as helping to
waterproof the ant's cuticle ('skin'). Certainly keeping clean and hygienic is especially important in a social animal
which lives in close proximity to others, facilitating the spread of disease. Licking one-another is also thought to
enable the ants to come into contact with odour exudates, so that any differences in individual body odours are
'smeared out' to create the odour unique to each colony which aids identification of ants belonging to the colony
and those that do not.
Ants also use their mandibles when carrying and digging. Digging may involve pawing movements of the forelegs,
sweeping soil particles back and underneath the body, but may also involve removing larger particles by lifting
and carrying them in the mandibles. The mandibles are also used to transport other ants. Usually the ant doing
the carrying grabs the other by holding one of its mandibles between its own mandibles. The ant being carried
tucks its head downwards with its body curled over and its legs held in and becomes immobilised, so it can be
carried as a rigid object.
Mandibles are also used by ants as weapons. Typically when two ants fight, the attacker will charge and grab the
other ant with its mandibles, often around a leg or antenna or mandible. The recipient ant may lose an
appendage or receive a puncture to its head capsule. Prior to such an attack, the ants may threaten
one-another: raising their heads with the antennae directed toward the enemy ant and the mandibles held wide
open with the labial mouthparts tightly withdrawn for their protection. The enemy ant once seized may be dragged
from the nest.
Another interesting feeding behaviour in these ants is food-sharing. One ant may solicit another to give it food,
or a well-fed ant may offer in the hope of inducing an ant to accept. The acceptor raises its head and forebody
and often forelegs off the ground with its antennae directed towards the donor's head or mouthaparts. Its
mandibles are closed. The acceptor laps up liquid food, regurgitated by the donor, with its hypopharynx, with its
maxillary palps held back. Meanwhile the donor also raises her head and forelegs but with her mandibles open
and her antennae passive. Her labial mouthparts and hypopharynx are extruded as a drop of regurgitated fluid
appears as the labial mouthparts move rhythmically. The ants may initiate this food-sharing behaviour by
touching one another with alternating forelegs on the cheeks or forelegs.
Another food-related behaviour of these ants is worth mentioning - gaping. This occurs in an ant that has fed for
a long time. The head is bent down and the mandibles opened. The labial mouthparts are extruded and a pellet
may extrude from the infra-buccal pouch (a pouch inside the oral cavity). This is thought to remove indigestible
particles filtered off during feeding and retained in the pouch.
This acts as a temporary food store when the midgut is filled to capacity. It stores mostly liquid food, whether
blood sucked up by a tsetse-fly (Glossina) or partially-digested food sucked up by a housefly (Musca). In the
mosquito, blood goes straight to the midgut, whilst fruit juices are stored in the crop (the labium has sweet
taste-sensors, the labrum blood taste-sensors). In the cockroach, Periplaneta, some digestion occurs in the crop
and here fatty acids (from the digestion of fats) are absorbed. The crop may contain still active salivary enzymes
and also enzymes refluxed forwards from the midgut. In bees, nectar is converted into honey, whilst in the crop,
by salivary enzymes.
The crop also has a function during molting in some insects, taking up air to expand the insect during shedding of
the cuticle and during enlargement of the insect prior to the new cuticle hardening.
Oesophageal Diverticula
Blind ending tubes or sacs may extend from the oesophagus; these are called diverticula. In saw-fly larvae
(Lophyrus) a pair of such pouches receives useless resin from the surface of pine needles on which it feeds. In
mosquitoes the diverticula take up any air sucked up with the food.
Function of the Proventriculus
This may simply be a sphincter or a muscular organ armed with spines or teeth, also ending in a sphincter. It
controls the passage of food into the midgut, acting like a sieve. In the cockroach, Periplaneta, a ring of six
powerful teeth break-up and compact the food. Behind these are two rows of hairy cushions. In the bee, the crop
filters out pollen grains that are passed onto the midgut, whilst retaining fluids such as nectar.
The Midgut
The mid-gut has muscular walls and is lined on the inside by cubical or columnar epithelium bearing microvilli or
some similar structure to increase the surface area for the exchange of materials with the gut contents. The
midgut lumen is not lined by cuticle, but is covered by the peritrophic membrane which protects the epithelium
and is secreted by the epithelium. This membrane may contain chitin, proteins and glycogen and is moulded by
elaborate mechanisms under the control of the gut wall musculature. It is periodically shed (and regenerated) and
encloses undigested food as it passes into the hindgut, forming a sheath for the faecal pellets.
The midgut secretes digestive enzymes and also absorbs many digested nutrients, such as glucose (the product
of carbohydrate digestion) and amino acids (the product of protein and polypeptide digestion). The fat body of
the insect converts absorbed glucose into trehalose, the sugar found in insect 'blood' (haemolymph) and body
fluids. Transporting sugar around the body in the form of trehalose makes it much harder for bacteria to
metabolise and this helps limit infection. (In mammals, the nervous system requires glucose and carrying glucose
in the warm blood necessitates a very advanced immune system in mammals). The rehalose is converted into
glucose for use when taken up by the cells.
The midgut may have distinct subdivisions. For example, in the tsetse fly (Glossina) the first region absorbs water
from the blood meal; the second region releases enzymes (causing the blood to blacken as it is digested); and
the final region absorbs the products of digestion. In some Homoptera, which feed on plant juices, the gut folds
around so that the oesophagus opens both into the front of the midgut, as usual, and also into a special filter
chamber in the back of the midgut. This allows excess fluid to short-circuit the digestive action of the midgut to
be excreted (the Malpighian tubules cover the surface of the filter chamber). Pouches or midgut caecae may
extend from the midgut (as in Aleochara at the top of the page) increasing its surface area for release of enzymes
or for absorption.
The Hindgut
A circular muscular valve, the pyloric sphincter, separates the midgut from the hindgut. The hindgut is lined by
cuticle often bearing tiny backward-pointing spines to help draw the faecal pellet backwards. The faecal pellet is
encloded in the shed peritrophic membrane of the midgut. The rear of the hindgut may form a distinct rectum.
This may bear six longitudinal folds or rectal pads or glands that may allow the rectum to expand when needed
and probably absorb water (the rectal pads are richly supplied by oxygen via tracheae and so are metabolically
active and the cuticle covering them is often thin and even porous). The rectum thus appears to act to conserve
water by absorbing water from the faeces. When you think how little water there is in a small insect, it is
remarkable how they can survive for so long between meals without drying up! A starved fly is thought to typically
run out of fuel (glucose/trehalose) before it dies by dehydration.
The faecal matter becomes progessively drier as it passes along the hindgut and ammonia and bicarbonate may
be secreted into it as the hindgut appears to regulate the pH (acidity) of the insect body. In the cockroach food
takes 9-33 hours to pass through the gut, in the silkworm 2-3 hours. Faeces are expelled from the rectum through
the anus, typically as dry pellets, except in those insects that have excess fluid in their diet, such as plant
sap-sucking Hemiptera, flies (Diptera), nectar-feeding moths and butterflies (Lepidoptera) and Hymenoptera
(bees, ants and wasps). In aphids and coccids (scale insects) it contains excess carbohydrates from the sugary
phloem sap of the host plant and is expelled as droplets of 'honey-dew' (often coated in wax in aphids to prevent
the aphid colony becoming covered in sticky honey-dew!). The beetle Platydema spins a pupal cocoon during
metamorphosis of chitinous fragments glued together, derived from the fungus on which it feeds.
Dietary Requirements and Symbionts
Insects are truly remarkable in their abilities to utilise unlikely food sources. They are also remarkable for the
efficiency with each they feed and grow, often on a tiny amount of food. Insects with distinct larval stages also
have the advantage of utilising different food sources, with the larvae avoiding competition with the adults by
eating very different food substances, such as the hungry caterpillar who eats foliage whereas the adult moth or
butterfly may feed on nectar. In many cases the larvae are aquatic and so feed in a very different environment to
the terrestrial adults. Almost anything organic is a food source for at least one species of insect. Cockroaches
can live on paper and a little water, silverfish may be kept on nothing more than a small piece of potato. House
flies, Musca domestica, can be grown using meat for their larvae (on which they will lay their eggs) and milk
powder and perhaps a little cholesterol, and water for the adults. The rove beetle, Aleochara bilineata, has a
high-protein diet of fly maggots, but can be reared happily on cat biscuits!
A good example to consider are the wood-boring insects. An example is Lyctus, the powderpost beetle, whose
larvae bore into wood, leaving behind powdery frass (faeces) of undigested wood. Wood is typically about 50%
cellulose ( a carbohydrate), 40% lignin and 10% hemicelluloses (another carbohydrate) and small amounts of
starch and other carbohydrates. Lyctus feeds on the latter - the starch and other sugars, but cannot digest the
main cellulose and lignin bulk of the wood which passes through the gut to be defecated as the powder which
gives the insect its common name.
Termites, however, have flagellated pro digest particles of wood and break-down the cellulose. The insect can
then digest excess protozoa and so derive nourishment. In this way termites will extract much of the cellulose from
ingested wood, but none of the lignin. Mixotricha is one of these protozoal endosymbionts found in the gut of
the termite Macrotermes (endosymbiont - an organism that lives inside another organism) and is a very unusual
single-celled organism indeed! Mixotricha uses its four flagella to steer when swimming, but locomotive power is
achieved by hundreds of thousands of spirochaete bacteria that live attached to the outside of the protozoan cell,
functioning as cilia, and drive it along (by means of their endoflagella). Spherical bacteria living inside the
protozoan cell provide it with energy by respiration, functioning as mitochondria which it lacks. Crucially, for the
termite, Mixotricha has at least two bacterial endosymbionts that can produce cellulase, the enzyme which
degrades cellulose into smaller carbohydrates, allowing it to degrade the particles of wood that it ingests. Thus,
the termite is a complex example of different organisms living together as a single unit! No animal is known to
synthesise its own cellulase, but to rely on bacteria to synthesise it for them. Cellulase is a bacterial invention! No
insect appears able to digest lignin, and this is defecated. (Some bacteria and fungi do digest lignin, often at a
very slow rate - lignin really is tough stuff!).
Many insects, if not all, rely on bacteria and other microorganisms living within their guts to assist in food
digestion. Vertebrates have a similar dependency; in herbivorous mammals bacteria in the gut secrete cellulose
to help digest plant cell walls and these live within special fermentation chambers, such as the appendix and
large intestine or a special additional compartment of the stomach, as in ruminants. The guts of many insects also
have special fermentation chambers. These chambers may be extensions or caecae of the midgut, such as the
bacterial crypts of Aphanus, which feeds on plant juices; or there may be a special chamber in the hindgut, such
as the rectal pouch of the termite Eutermes, which contains flagellates, or the fermentation chamber of the
lamellicorn larva. In humans the bacteria inside in the gut lumen, as they do in many insects, but in many insects
the symbionts are often intracellular - living inside the cells, often special cells in the gut lining.
Humans acquire their bacteria from the environment and from their diet, but in many insects the microorganisms
are very specific and it may be crucial that the insect acquires its special bacterial symbionts. In these cases the
mother often passes them on to the offspring. In the bed bug Cimex, the symbionts enter nurse cells which
surround and nourish the developing egg in the ovary and pass from here into the egg cell itself. In a similar
manner rod-shaped bacteria living in the fat body of the termite Macrotermes are transmitted to the egg cell whilst
in the ovary. A similar invasion of the egg, but just before the egg shell is secreted, occurs in many insects, and
Lyctus (powderpost beetle) mentioned earlier. In some insects the bacteria infect the male reproductive system
and are then passed onto the male and enter the egg through the micropyle before it is laid (the micropyle is a
pore in the shell through which the sperm enters during fertilisation). There may be special glands attached to the
male reproductive system that store and release the microbes for this purpose. In some the female contaminates
the eggs when they are laid and the hatchlings eat all or part of the egg-shell, or probe its surface, to pick up
their microbial partners. The symbionts may be bacteria, protozoa or fungi.
Other uses of the mouthparts
Some insects have evolved diverse uses for their mouthparts. Ants are a good example and the behaviour
patterns of Formica fusca have been well studied by D. I. Wlllis (Behavioural patterns of the ant, Formica fusca,
1962. Animal Behaviour 10: 105-111). The ant Formic fusca feeds on liquid foods by lapping with its 'tongue': a
bulbous projection, properly called the hypopharynx, on the labium (lower lip). During this ingestion of fluids the
mandibles are closed and maxillary palps are held back out of the way. The antennae are held in a W shape with
their tips resting on the surface of the food along with the labial palps - both antennae and labial palps seem to
be involved in tasting the food.
These ants also use their mouthparts for cleaning. When cleaning the antennae and forelegs, the fore-part of the
body is lifted 30 degrees above the horizontal, so that the middle and hindlegs are left supporting the body. Left
and right appendages are cleaned alternately. For example, when cleaning the right antenna, the right foreleg is
wiped down the antenna which is drawn through the strigil (a comblike spur of more-or-less rigid hairs) of the
foreleg as the head is held inclined towards the side being cleaned. The antenna is wiped one to three times in
succession whilst simultaneously the left foreleg is drawn through the mouthparts on the left side, between the
maxilla and labium to be cleaned by the maxillary comb (and possibly the hypopharynx). Sometimes, as part of the
same sweeping movement, the right foreleg may begin by sweeping over the back of the head and eye to clean
these structures too. The mandibles may also be brushed downwards by the left foreleg before it is drawn
through the mouthparts. This pattern is reversed as the left antenna and right foreleg are cleaned and this
alternating pattern is repeated several times.
An alternative posture is adopted when cleaning the middle and hindlegs. This time the ant leans away from the
side being cleaned and raises the legs on the side being cleaned whilst its body is supported on the middle and
hindlegs of the opposite side and the abdomen (forming a stable tripod). When cleaning the left legs, for example,
the left hindleg is cleaned by the left midleg brushing down it (which maybe assisted by the forelegs). The left
midleg is then cleaned by both forelegs brushing down it in succession. The tarsi ('feet') are cleaned mainly. Thus
there is a sequence in which foreign particles are passed from the hindlegs to the middle legs to the forelegs and
then to the mouthparts. Forelegs may also be brushed down each other as one leg is pulled through the strigil of
the other. Hindlegs may also brush the side of the abdomen before being cleaned.
Abdominal cleaning utilises a third posture. The body is raised and held clear of the ground by all six legs as the
head bends underneath the thorax whilst the abdomen is bent forwards towards the head. The mouthparts then
lick the end of the abdomen. The ants may also lick one-another, coating each other with saliva and cleaning
parts that the ant cannot reach by itself. Mouthparts, the abdomen and limb joints are especially focused on. The
ant being cleaned remains passive as the cleaning ant plays its antennae over the surface of the recipient.
Wallis speculated that the oily saliva of the ant may contain anti-microbial properties, as well as helping to
waterproof the ant's cuticle ('skin'). Certainly keeping clean and hygienic is especially important in a social animal
which lives in close proximity to others, facilitating the spread of disease. Licking one-another is also thought to
enable the ants to come into contact with odour exudates, so that any differences in individual body odours are
'smeared out' to create the odour unique to each colony which aids identification of ants belonging to the colony
and those that do not.
Ants also use their mandibles when carrying and digging. Digging may involve pawing movements of the forelegs,
sweeping soil particles back and underneath the body, but may also involve removing larger particles by lifting
and carrying them in the mandibles. The mandibles are also used to transport other ants. Usually the ant doing
the carrying grabs the other by holding one of its mandibles between its own mandibles. The ant being carried
tucks its head downwards with its body curled over and its legs held in and becomes immobilised, so it can be
carried as a rigid object.
Mandibles are also used by ants as weapons. Typically when two ants fight, the attacker will charge and grab the
other ant with its mandibles, often around a leg or antenna or mandible. The recipient ant may lose an
appendage or receive a puncture to its head capsule. Prior to such an attack, the ants may threaten
one-another: raising their heads with the antennae directed toward the enemy ant and the mandibles held wide
open with the labial mouthparts tightly withdrawn for their protection. The enemy ant once seized may be dragged
from the nest.
Another interesting feeding behaviour in these ants is food-sharing. One ant may solicit another to give it food,
or a well-fed ant may offer in the hope of inducing an ant to accept. The acceptor raises its head and forebody
and often forelegs off the ground with its antennae directed towards the donor's head or mouthaparts. Its
mandibles are closed. The acceptor laps up liquid food, regurgitated by the donor, with its hypopharynx, with its
maxillary palps held back. Meanwhile the donor also raises her head and forelegs but with her mandibles open
and her antennae passive. Her labial mouthparts and hypopharynx are extruded as a drop of regurgitated fluid
appears as the labial mouthparts move rhythmically. The ants may initiate this food-sharing behaviour by
touching one another with alternating forelegs on the cheeks or forelegs.
Another food-related behaviour of these ants is worth mentioning - gaping. This occurs in an ant that has fed for
a long time. The head is bent down and the mandibles opened. The labial mouthparts are extruded and a pellet
may extrude from the infra-buccal pouch (a pouch inside the oral cavity). This is thought to remove indigestible
particles filtered off during feeding and retained in the pouch.